
技术交流

扫描二维码
或添加“GeneGroup003”
获取更多更新资讯
商城订购

扫描二维码
或添加“基因商城(GeneMart)”
手机下单,快人一步
售后服务

扫描二维码
或添加“GeneGroup005”
获取更快速售后支持
买2送1--坏死性细胞死亡信号通路抗体推荐

细胞坏死由显著的化学或物理损伤所引起,曾被定义为是被动和非程序性的细胞死亡形式。近期,研究数据显示坏死性细胞死亡也可以通过不依赖 caspase 的活跃的程序性过程发生,这种坏死形式称为坏死性凋亡,可被细胞外信号(死亡受体-配体结合)或细胞内诱因(微生物核酸)所诱导,并且受到 caspase 活性的显著抑制。此外,另一种由炎症 caspase 蛋白介导的程序性坏死称为焦亡,焦亡在天然免疫应答中发挥重要作用。
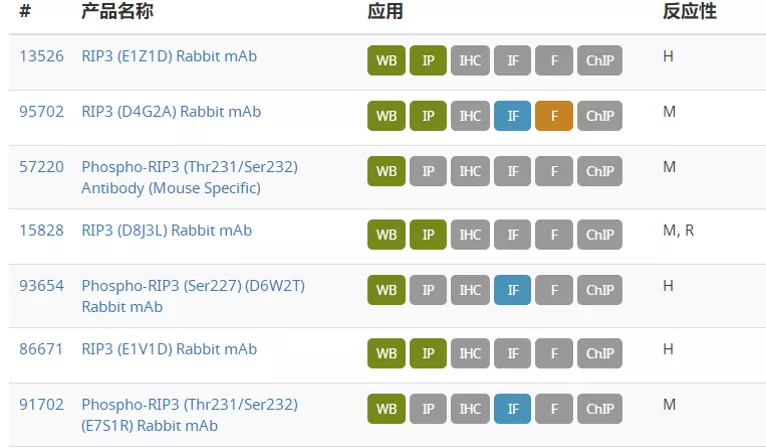
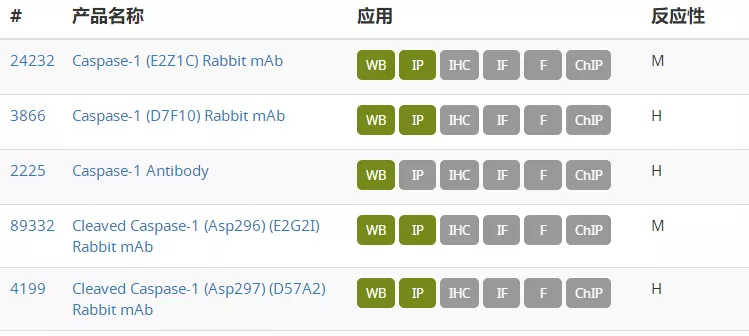
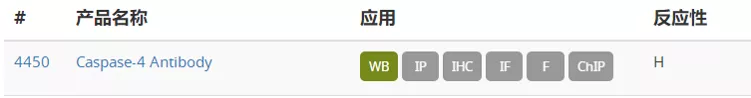
坏死性细胞死亡信号通路图 图片来自CST(Cell Signaling Technology) 研究显示,坏死性凋亡在癌症以及几种神经退行性疾病中发挥重要作用。坏死性凋亡可以参与肿瘤转移,抑制坏死性凋亡通路则能够限制肿瘤生长。存在 necrostatin-1 的情况下坏死性凋亡会受到抑制,据报道在阿尔茨海默病和帕金森病中,Necrostatin-1 处理能够提高细胞活力。另外,关于靶向焦亡的治疗可能性的研究提示,焦亡对于癌症、自身免疫性疾病和神经退行性疾病以及其他病理状态有益。有研究证明 ω-3 脂肪酸在三阴性乳腺癌细胞中能激活焦亡,表明焦亡具有介导癌症的潜力。因此,研究坏死性凋亡及其他细胞死亡通路的机制可为各种疾病的新型治疗方式开发提供具有治疗价值的见解。 今天小编就整理一些坏死性细胞死亡的关键靶点抗体供大家参考。 RIPK1、RIPK3 和 MLKL 是参与坏死性凋亡的关键蛋白。RIPK1具有激酶活性,是多种复合物的重要组成部分。Ser166、Ser161 和 Ser14/15是RIPK1 的自磷酸化位点,这些位点可被 Neec1 抑制。IAP 会引起 RIPK1 泛素化,导致RIPK1 在 Ser320 位点的磷酸化, NF-κB 激活及后续的炎症级联。RIPK1可与另一个激酶RIPK3 结合形成程序性坏死复合物IIb。在坏死性凋亡期间,RIPK3 在 Ser227 位点被磷酸化,此磷酸化为 MLKL 激活所必需,MLKL 作为 RIPK1 和 RIPK3 下游的效应蛋白发挥作用。RIPK3在Ser358位磷酸化MLKL,导致其寡聚化和转运到质膜并行成孔道,最终导致细胞坏死和死亡。 在典型的细胞焦亡过程中,由胞质 PRR、接头蛋白 (ASC) 和 pro-caspase-1组成的炎症小体组装后,pro-caspase-1 被蛋白水解激活并将细胞因子 pro-IL1β 和 pro-IL18 剪切为其成熟的促炎症形式,即 IL1β 和 IL18,并诱导 gasdermin D 的剪切。细胞内的胞质 LPS 诱导非典型焦亡,导致 gasdermin D被活化 的caspase 4*、5* 和/或 11 剪切。Gasdermin D (GSDMD) 是炎性半胱天冬酶的一种底物,经剪切会产生两种片段:GSDMD-N 和 GSDMD-C。Gasdermin D的剪切在典型和非典型焦亡激活中均发挥作, Gasdermin家族的成员GSDMA、GSDMB 和 GSMDC也已成为细胞焦亡的关键组分。 参考文献: Couillin, I et al. (2011) 978-3-0348-0148-5. 开学促销正在火热进行中!2021 年3 月 1 日至 5 月 31 日,一次性购买 2 支 S 或 L 包装一抗, 即可免费获得 1 支Microsize(T包装)一抗,活动详情请戳下方链接 基因有限公司作为CellSignalingTechnology (CST) 优质一级代理商,为您实验提供更优的支持和帮助。这些坏死性细胞死亡的关键靶点抗体在促销之列,赶快抓住机会,联系基因有限公司人员来参加吧!想要更多相关知识和资料,或了解更多促销信息请在本文后留言或联系您身边的基因有限公司员工。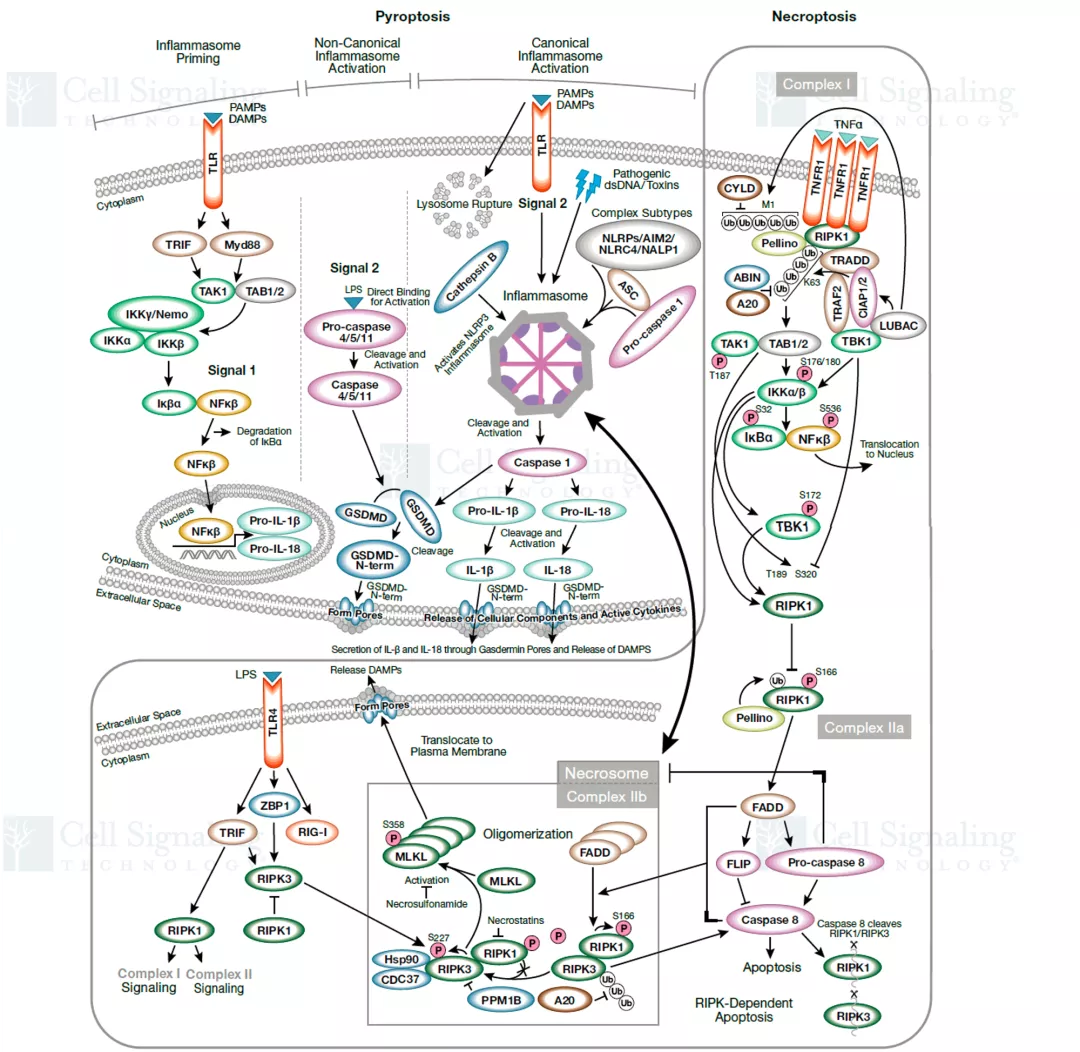


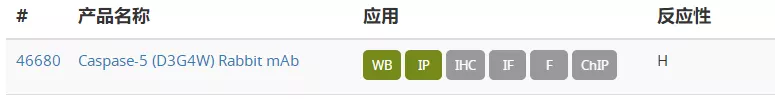
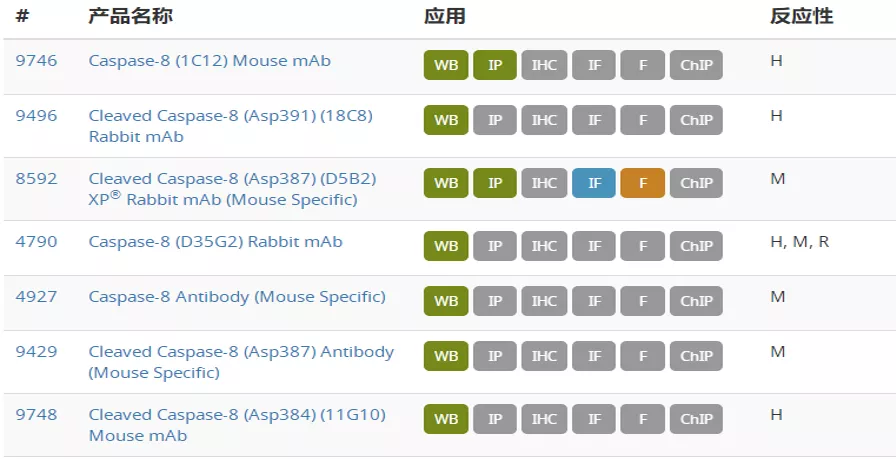
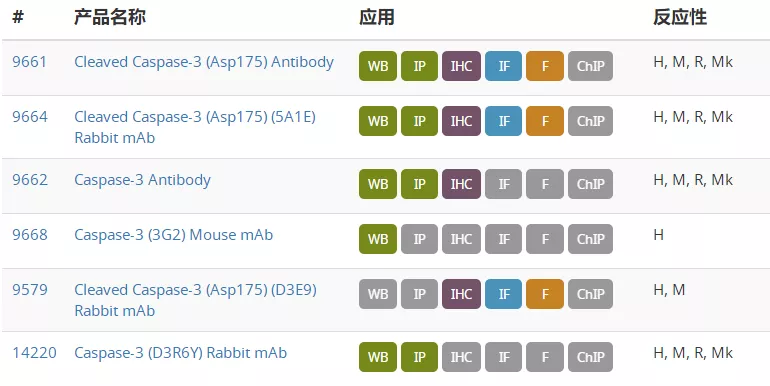


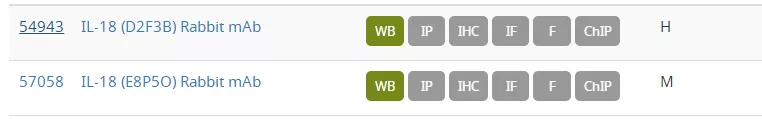
de Gassart A, Martinon F (2015) 43(5), 835–7.
Festjens N et al. (2006) 1757(9-10), 1371–87.
Galluzzi L, et al. (2017) 12, 103–130.
Guo H, et al. (2015) 21(7), 677–87.
Gurung P, et al. (2015) 16(6), 596–7.
Lim Y, Kumar S (2015) 6(35), 36926–7.
Martin SJ, Henry CM (2013) 61(2), 87–9.
Petersen SL, et al. (2015) 22(11), 1846–57.
Ramos-Junior ES, Morandini AC (2017) 40(6), 313–316.












